#speculative biology

The Early Temperocene: 145 million years post-establishment

The Map and the World of the Early Temperocene
The Temperocene is a far different world than the Glaciocene that came before. Melting ice and rising sea levels have flooded most of the globe, to an extent that 80% of the surface is now water. The continents have significantly shrunk, their ragged edges and offshore isles a telltale sign of land swallowed by the sea, while meanwhile in the ocean, vast swathes of shallow sea, breeding grounds of kelp forests, coral reefs and seagrass mats scatter in abundance.
TheCentralic Ocean is now but virtually gone: now divided into multiple small seas connected by narrow straits. Ocean currents cycle in loops around the continents, creating a turbulent pattern keeping the seas cycling cold and warm water, producing, in turn, warm and cold sea breezes that affect the climate even on the air and land, creating random, irregular patches of temperate forest and tropical jungle all throughout the continents due to the varying temperatures and the plentiful rains. These currents also cycle nutrients from the deep back to the surface, providing sustenance to sea plants and phytoplankton, two important food sources and the basis of all of the ocean’s food webs.
Scarcely any ice caps remain, except on the very northmost and southmost tips of the continents. The Glaciocene continents still remain somewhat similar, though with a few important changes. South EasaterraandPeninsulaustra have since merged into the continent Australo-Easaterra, and have traded their fauna: blubbats and blubbears have moved into the plains of Easaterra, while carnivorous rhinocheirids and tetracorns have migrated into the Peninsulaustran continent. Isla Centralis has fused with Mesoterra, and despite a rough start, its native animals– the rabbeasts and the scabbers– are flourishing alongside Mesoterran locals such as the podotheres and the piggalo.
But the most significant effect of the Temperocene is the formation of isolated landmasses: as sea levels rose, islands were cut off from the mainlands: and so were the flora and fauna from their relatives. Exposed to new environments, they begin to evolve separately, giving rise to highly-divergent forms. On North Westerna, relics of the hammoth lineage, shrunken by time, now have to contend with a new predator from the skies. On the offshore island of Isla de Oof, a beast of hulking inefficiency soldiers on in spite of everything. On the North IslesandIsla Junctus, the last fragments of tundra linger on from an age of ice. On the new subcontinent of Fragmus, lineages once marginal and insignificant find new opportunities for diversity and greatness. And in the temperate forests of South Ecatoria, thirty million years after the demise of the harmsters, a promising new species–or two– glimmer with the potential of a new intelligence.
The Temperocene has brought about a diversity unlike anything that the history of the planet has ever seen before. Numerous small clades, marginal in the harsh climates of the Glaciocene, now find perfect opportunities to spread, adapt and evolve. Now, land, sea and sky teem with stunning abundance and difference: an array of various forms that will only grow as the forces of natural selection shape the species to their different environments.
▪▪▪▪▪▪▪▪▪▪▪▪▪
The Early Temperocene: 140 million years post-establishment

Dare To Be Stupid: The Big Oof
The favorable conditions of the Temperocene have seen evolution working wonders in ascending life to a degree of diversity and derivedness like never before. Thousands, if not millions, of new species have emerged to take advantage of so many new niches available in the increased warmth of the globe: and even more so following a mass extinction whose vacant gaps have quickly been filled by new pioneers.
With increased competition giving rise to ever-more efficient grazers, hunters, swimmers and flyers, an illusion of progress is made prevalent, as natural selection weeds out the poorly-suited ones unable to compete for a niche in the environment. Yet, on one small island off the coast of Arcuterra, one very unusual animal, remarkable for seemingly all the wrong reasons, challenges this very notion, a living testament to evolution’s lack of a direction, and the complete absence of any goal except current survival. It is a highly aberrant member of the lemunkies: a clade of social, intelligent and adaptable creatures, with some possibly even bordering on semi-sophonce– yet this beast is anything but.
Known as the big oof(Tardipithecus stultus), this lumbering browser is everything that its relatives are not: a slow-moving, clumsy, solitary animal that does little else but spend much of the day highly inactive, with intermittent periods of constant feeding, on tough, scarcely-nutritious and highly-toxic foliage. Doing little else other than pluck leaves from branches and stuff them in tremendous quantities into its mouth, the big oof has thus abandoned one of the lemunkies’ most valuable assets: their large and adaptable brains. The big oof, despite standing almost six feet on its hind legs and weighing as much as five hundred pounds, has a brain scarcely larger than a potato and weighing little more than half a pound: further confounded by its smooth texture lacking most of the folds and wrinkles typical of the mammalian brain. This adaptation gives the big oof a very distinctive flat-headed profile, and extremely inflexible behaviors: its diet of toxic plants, primarily weedwood pseudo-trees, means it just barely gets by on just enough energy to survive, and as it feeds by reaching up to tear branches from trees, if it sees any leaves on the ground it will no longer even recognize it as food: as such the big oof is a very messy eater, as any food it drops from its grasp may as well have ceased to exist in its perspective.
This complete lack of brainpower and utter lethargy makes the big oof a highly-solitary creature, unlike its gregarious close living relatives, the chimpmunks and the hamrambes. In fact, the big oof basically even lacks any capacity to recognize or interact with members of its own species, simply passing them by should two meet by chance, and any semblance of social interaction occurs only during mating, when males are attracted by the scent of a receptive female, easily the only time any thought other than food ever crosses their tiny, simple brains. There is no courtship, no affection, no competition, or even anything resembling pair-bonding: the male simply mounts the female and instinctively completes the act in but a few minutes, while she is almost entirely oblivious to his presence, and once the job is done, he departs with no further interest. The female then gestates a single offspring for as long as a year, due to her nutrient-poor diet, before delivering a well-developed pup that can walk nearly right away. The complete lack of predators in their island means that mother oofs barely even bond with their young, simply passively allowing them to nurse on the thin, watery milk, and once weaned, at around a year and a half due to the low-fat content of the milk, again from the mother’s poor diet, the youngster simply wanders off to forage independently– and the mother scarcely notices the absence and forages as she always has. Instinctively, the youngster learns with no guidance how to pull off branches and shovel them into its mouth: about the only thing a big oof even knows in the first place.
The big oof thus seems like a very terribly designed creature, and indeed it may well be. Slow, lethargic, dim-witted to an extreme, virtually defenseless and, worst of all, completely incapable of adapting to new behaviors, it looks and acts every bit like an utter evolutionary failure inherently doomed to extinction. Yet, despite all, the big oof is, quite unbelievably, a triumph of natural selection. Stranded on an island with no danger, but also little other food except the abundant but highly-non-nutritive weedwood, the ancestors of the big oof were shaped, generation after generation, to become suited to this harsh new environment. An absence of useful calories made a large and energy-hungry brain a liability, and so over time its brain atrophied to such a level as to just remain functional enough for the animal to live. There were no predators on the island, so the oof lost all fight-or-flight responses or protectiveness toward its offspring. There was no benefit to social groups, so it lost its higher capacity to recognize and empathize with its own species. And as there was an abundance of one particular food with weedwood regrowing very fast due to its rhizome system and numerous shoots per plant, energy and resources was better off spent on a digestive system that can efficiently squeeze every last caloric drop from this one food source, at the expense of basically every other bodily function: it didn’t need to be agile, to move fast, or to be flexible or creative in finding its food– all it had to do was reach up, pluck, and stuff in its mouth, reach up, pluck, and stuff in its mouth…rinse and repeat, every single day, for the rest of its 30 to 40-year life.
Perhaps, indeed, the big oof has effectively doomed itself by crippling overspecialization. If faced with new food sources, it would simply ignore them, their survival benefits all but alien to its smooth little brain. If threatened by new predators, it would not understand the danger they pose to itself or its young. If challenged by even the slightest change to its secure environment, the big oof would literally just sit in place until it starved to death. And yet, by thriving on a food source that would be useless to any other creature, by surviving in a place where most other large animals would struggle to find enough nourishment to fuel their bodies, the big oof, nonetheless, is an evolutionary success story. It illustrates as a perfect example of evolution’s lack of foresight, not geared toward making organisms “better”. For perhaps tomorrow, or in a million years, the specialized big oof might be faced with changes it cannot adapt to and go extinct. Yet today, it thrives in the environment it adapted specifically to: hanging on not as a demonstration to the survival of the fittest, or survival of the strongest or the smartest: but the survival of the simply good enough.
▪▪▪▪▪▪▪▪▪▪
The Early Temperocene: 140 million years post-establishment

Phyte or Flight: Flora of the Early Temperocene
The Temperocene Era has seen the diversification of animal clades that re-emerged after the end-Glaciocene mass extinction, filling ecological niches left vacant by the victims of the catastrophe. But animals are not the only things that evolve– it is easy to forget that plants too, despite their seeming inert nature, are equally struggling in the game of life in a quest to rise to the top: fighting, competing, and migrating just as animals do, albeit on a far slower timescale.
The Temperocene, thus, is lush with new plantlife, the warm and humid clime favorable for the survival of dense temperate forest and tropical jungle, as well as vast grasslands, steppes and scrubland home to millions of species of flora that stabilize the food web from the bottom up.
Most influential, of course are the trees: conifers, once abundant in the Glaciocene, have now receded further south and north, and decidious trees, the stonefruits and citrus, have returned to much of their former range in the Therocene. While most of them have remained generally-unremarkable variants of the basal state, a few odd specialists have taken hold of unusual new niches suiting a warm tropical clime.
Coastal beachpeach(Littoraprunus dispersus) is one of the most successful trees in the Temperocene. Adapted to coastal marshes and resistant to saltwater and submersion, this stonefruit tree stands on stiltlike roots to allow it to breathe, lifting its main trunk well above high tide line. Its buoyant fruits are able to migrate long distances, floating on ocean currents and taking root on isolated islands: and while it originated on the south of Arcuterra at the Glaciocene’s end, this highly-enterprising fruit of the sea has now made home on nearly every coast: even the northern, non-frozen coastline of icy Peninsulaustra.
Another unique tree of this age is the dwarf desertade(Aridocitron succulens), a citrus species thriving in another biome more common in the hot age: arid desert. Where once chilly drylands spread, now the heat has turned up, spelling doom for many cold-desert plants and the animals that eat them. Yet the desertade thrives. Its trunk, short and stocky, contains spongy tissue that store water, while thick waxy leaves prevent dehydration and thorny branches deter herbivores. But while herbivores are a bane to the desertade, it does enlist their help for a crucial point of spreading its seeds. And there is no better way to draw in thirsty travelers than with a refreshing drink: thus, the desertade’s fruit are big, juicy and are less sour than most of the citrus family, a welcome trade for desert animals in the price of transporting its future generations.

Strangely, however, plenty of what appear to be trees are in fact not trees at all. Indeed, even on Earth, the classification of “tree” is a polyphyletic one, with numerous unrelated plants becoming large, rigid towering giants. And here on HP-02017, several plant groups would follow the trend: though functioning quite different from a proper tree.
One such example is the grovegrass(Arbomimogramen spp.), a type of weedwood growing up to eight feet in height. Its kind first grew in such heights following the extinction of their primary consumer, the mison, at the beginning of the Glaciocene. With the tropical Temperocene weather conducive to their growth, grovegrass quickly spread to most continents by its floating dandelion-like seeds, where it now grows as the understory of forests, and at times even dominates large-plant niches solely on isolated islands.
And it is not the only grass to mimic a tree: the notorious saberleaf, in northern region of Gestaltia that was once Borealia, now grows as the twelve-foot sabertree(Arbogladius spp.), largest of a subclade known as sabershrubs and every bit as sharp and abrasive as its ancestor– yet a favored food of the tall ungulopes known as altolopes, which had replaced the now-extinct girats. Another towering grass, the spiny bramboo(Spinaschoenus spp.), also forms prickly forests reaching as high as twenty feet in tropical regions across Gestaltia, where specialized unicones and badgebears feed on their abundant growths.
One major advantage these tree-mimics would have over true trees is that they were, despite their greater size, still grasses: growing from a singular underground rhizome and sending up shoots as they spread. As many as a hundred separate “trees” are thus just a single plant, allowing them to regrow quickly and sprout up new shoots: making them a resilient and abundant food source that can sustain even large quantities of browsing herbivores, all too common in the Temperocene.
Other, unrelated plants too would reach tree-like proportions. Sandfans(Monofoliopluma spp.) are towering cloverferns adapted for desert life, filling a role akin to cacti with their thick sturdy stem and leaves fused into a single giant plume that minimizes water loss in dry climates. Also abundant are cabbage descendants, the brasstics, which grow into huge, herbaceous mock-trees, such as the swamp-dwelling tropical salaspire (Brassicarbor spp.), and the mountain-growing, cold-adapted mountplume(Montanobrassica spp.), which is a vital food spurce for many mountain dwellers such as ungulopes and cragspringers.

Smaller plants, of course, are vastly more abundant: forming low-ground foliage the carpets plains and steppes and serves as plentiful forage for grazers. Ever present are more-typical grasses, such as the prickly thornbriars(Gramenosentis spp.), and the Gestaltian flameweed(Pyrogramen spp.) with its stinging chemical defenses few herbivores can tolerate. These grasses are abundant as ever as they were prior to the Glaciocene freeze, and today are vastly diverse: each species with unique defences that favor niche partitioning among grazers.
Another extremely successful plant are the descendants of clover: initially introduced during planetary establishment as a forage crop, the plant has since flourished and filled many niches of herbaceous plants of virtually nearly every shape and size. Some are similar to the ancestral form, such as goldtip trefoil (Prototrifolium aureum), but others developed bigger leaves with more leaflets, giving rise to the cloverferns like the pinkpom cloverfern (Polyfolium rosae), which often spread across the forest floor. Some cloverferns have adopted unusual traits in the ever-enterprising quest to expand, such as coalclove(Erythrotrifolium trichromus), with distinctive red coloration due to having such quantities of red and black pigment as to almost mask the still-present chlorophyll, partly as a warning to grazers of its distasteful flavor, and partly to harness Beta’s faint rays during Beta-twilight.
Cloverferns, however, have long branched out into many diverse niches in the race for sunlight. Some, such as clovervines(Vitifolium spp.), have taken advantage of nearby woody plants as support to grow taller than any typical herbaceous plant, anchoring itself with grasping tendrils modified from branching stems. Others, such as torchbush(Foliofrutex spp.) become shrub-like bushes with more rigid stems and feeding low-level browsers on scrubland and forest floors. Some cloverferns have even taken to the water, like pondspinners(Aquatrifolium spp.), with large leaves floating on the surface and flowers on stalks that breach the surface during suitable breeding conditions.

The flooded world of the Temperocene, in fact, is highly conducive to aquatic plants, and many have not simply taken to rivers and lakes but to the sea: and are primarily responsible for the sheer diversity of herbivorous aquatic megafauna, a niche rare on Earth but extremely prevalent here. Primarily, grasses dominate the seas, alongside kelp and algae (which technically are not true plants), descended mostly from two varieties: the floating coast kudzu and the rooted undergrass.
Coast kudzu has reached peak diversity, forming blankets of vegetation on the surface that harbor mini-ecosystems of their own. The highly-prolific floatvine(Pelagogramen spp.) grows massive branching rhizomes extremely fast as to cover massive areas of surface water in the span of a few days, kept only in check by the grazing of hamatees such as whaleruses, its sheer abundance being key to support such enormous marine herbivores. Other coast kudzu, however, are far less spreading: the thick raftroot(Mareradix spp.) consists mostly of a single thick nutrient-storing rhizome, allowing it to thrive in colder seas and remain dormant until conditions are right, or the plantform(Pulpitophyton spp.), which forms small but dense mats of stems and roots: dense enough that small, sea-going ratbats occasionally roost on them, and also have been instrumental in allowing terrestrial minifauna, like duskmice, furbils, rattiles and various invertebrates, to raft from continent to continent and reach isolated islands, whenever such animals washed out to sea by accident or intentionally hitched a ride.
The other clade of marine grasses, the bottom-growing undergrass, in the meantime is mimicking the dynamics found on dry land by developing specific defenses to ward off herbivores: but, more often than not, ends up favoring the evolution of specialized herbivores that specifically evolve to breach their defenses. Whiteblot(Albomarinogramen detestus) is one such species, bearing distasteful compounds that it advertises with white markings easily visible in the blue light of the sea, and thornocopia(Spinatomar aquasentis) possesses spiny stems and leaves to deter herbivores from eating it. However, over time, sea grazers gradually develop resistances to such methods of defense, and a familiar trend begins to emerge: niche partirioning, rotational grazing, and specialized herbivores adapted to tackle one defense but not the other– a dynamic nigh-identical to the plains grazers on dry land.

Pressured by herbivore “predation”, competition with neighbors and the constant struggle to beat rival neighbors in the quest for water and sunlight, the merciless brutality of nature quickly surfaces in the plant kingdom. Nature red in tooth and claw is most evident in animals, visibly devouring and hunting one another in exuberant displays of the battle for survival. But plants are no exception. They too are living creatures simply operating at a slower frame of time, seemingly inanimate in an animal’s perspective. But they, too, fight one another, compete with one another– and at times, even actively try to kill one another and the animals that eat them.
One of the more blatantly vicious examples is the mace-bud destrotor(Vehemephyton telumgermen), a grass descended from the now-less common bleedweed. Stalk upon stalk, it rises along the ground as its rhizome steadily creeps sideways beneath the surface. But unlike its now-increasingly-rare relatives, the destrotor does not take kindly to competition. As each new stalk rises, it emerges as a spiral, with a tip covered in pointed hooked barbs, and once grown, the stalk gradually unfurls, spinning its barbed tip in circles like a mace in slow motion. With each spin, lasting about a day, it tears apart, uproots and impales neighboring plants within an eight-inch radius, purposefully killing off any competition for sunlight and soil nutrients and creating ominous, empty circles of barren soil around each new stalk that rises from the soil.
Some plants, in fact, not only eradicate their neighbors, but actively prey upon them. The white witherer (Radixavenator vampyrium) is a cloverfern growing along damp shady places like the forest floor, where, while seemingly innocent and even beautiful with its pale pinkish-white leaves, beneath the ground is up to something far more insidious. Its roots snake out long distances in search of other plants’ roots, and upon detection, latch onto them, pierce their outer layer, and release smaller rootlets to grow inside the host tissues and begin draining them of nutrients. So dedicated is the white witherer to attacking other plants that it has lost nearly all its chlorophyll, instead getting all its energy from stolen sustenance from other, photosynthesizing plants, which turn yellow and droop all around while it flourishes. Ultimately, the white witherer eventually runs out of neighbors to feed on, and as energy dwindles, the mature plant blooms, spreads its seed, and dies, leaving its next generation to continue its silent, botanical massacre.
Not only other plants are the victims of the Temperocene’s ferocious flora: animals too, particularly herbivores, are the main enemy of most plants, which try to defend themselves in many different ways from the eager gnawing teeth of rodent grazers big and small alike. Some sequester bad-tasting chemicals, develop hard outer coats, or produce thorns that make them painful to consume. But due to extreme herbivore pressure in its native range, one member of the weedwood family, the stinging barbriar (Mortiphyton vireum), takes its arsenal to a frightening new level. Coated with long, yellow spines, it secretes from the root of each one a potent cocktail of enzymes that, if injected into any would-be consumer, acts as a potent cytotoxin that causes cell death, internal bleeding, organ failure and even tissue liquefaction in minutes, with the barbed spines easily breaking off and remaining embedded in the target to continue delivering their deadly dose. Larger herbivores are lucky enough to leave with grisly swollen scars at worst, but small grazers, such as small hare-like hamtelopes, can easily acquire a lethal dose in as little as three or four spines, collapsing within minutes in violent spasms as they foam at the mouth and bleed profusely from every orfice as hemotoxic effects rupture their blood vessels– victims of the planet’s first venomous plant.
And while the barbriar simply targets animals in defense, other, more enterprising plants attack small creatures as a source of nutrition, especially in poor soils lacking in nitrogen– they have actively become carnivorous, trapping insects and digesting them to acquire vital and essential compounds otherwise lacking in their environment. A grass known as the shmuckbait(Flosacaptionem spp.) is a key example: native to nitrogen-poor deserts, it spreads its seeds and pollen by wind, and thus has no need for pollinators. Yet still, it produces colorful flowers with sweet-smelling nectar for an entirely different purpose: hungry bugs drawn by the scent and signals home in to feed– only to find themselves on the menu as they get stuck onto the sticky petals that then close over them to begin digestion.
Carnivory in plants, however, sometimes requires a little help, and a more indirect means of predation. The gluefern(Glutinofolium spp.) is a marshland cloverfern that, like the shmuckbait, produces sticky secretions to trap prey: but rather than emitting a sweet fragrance to lure pollinators, it instead emits the pungent smell of rotting meat to draw in scavengers such as carrion flies. But the gluefern cannot digest them on its own, as it lacks the necessary enzymes: instead, it enlists the symbiotic services of a specialized beetle: the gluefern bootbug (Decipulahabitus defecatum). The bootbug, equipped with specialized hairs on its feet and antennae and a waxy coating on its wing cases, is able to scurry about the gluefern without getting stuck: and feeds on the unlucky insects that get trapped in the plant. The plant basically functions as both home and prey trap, much like a spider’s web, and in return, the beetle’s droppings are a source of nitrogen compounds, now pre-digested and packaged for consumption. Thus plays out a peculiar case of mutualism: the plant acts as a home for the beetle that provides it with easy food– and all the beetle needs in turn is to pay its rent with poop.
The Temperocene has proven itself to be a lush haven for animal life to rapidly diversify into complex ecological webs, as unusual adaptations create ever more complicated interspecies relationships. Yet, among a world of increasingly unusual beasts, one must not forget the living things that make all them possible: living things that, despite appearing as mere scenery, are every bit as alive as their faunal counterparts: and every bit subject to evolution in all its beautiful, bizarre, and brutal ways.
▪▪▪▪▪▪▪▪▪▪▪
The Early Temperocene: 140 million years post-establishment

Not Quite Flight: Arboreal Gliders of the Temperocene
Since the Middle Rodentocene, the skies of HP-02017 had been filled with ratbats, quadrupedal flyers of distant relation to the squizzels and lemunkies, that had dominated the air for eons. And much more recently, they were joined in the air by the pterodents: airborne podotheres that, akin to the ratbats, would emerge from gliders, before truly taking to the skies on their own power.
That is not to say, however, that gliding was a prerequisite to true flight, as, in the Temperocene, several small lineages convergently had developed this form of half-flight– and stayed there. Evolution, being a random process with no goal except survival, allowed these seemingly-transitional forms to become a niche all their own, and as long as this partial airborne ability allowed them to thrive in their environment there was little need to progress further: unless, by chance, changing ecosystems or unexploited niches goaded them in the right direction.
One such species is the eyespot parachu (Volaticosciurumys oculus), a gliding kiterat descended from the lineage that would give rise to the ratbats: unlike its relatives, however, it would only use this ability to move from tree to tree and not much further. This was an easy way to traverse the forest, where the parachu could easily seek out its favorite food– insects in bark– as well as the sugary tree sap it spills while gnawing for bugs, supplementing its protein-rich diet with carbohydrates. The parachu thus is well suited for its current niche, and thus, unlike its ratbat cousins, has not evolved its patagia into a proper wing. The patagia, while used for gilding, serves another practical purpose: it bears large spot markings resembling false eyes, which it uses in defense: when startled by a predator, it spreads its patagia instantly, creating the illusion of a larger and more deadly creature, halting the predator in shock and giving the parachu time to glide to safety.
While the parachu glides with membranes of furred skin, stretched between its ankles and wrists, another common arboreal glider uses entirely different structures to get airborne: the plumetail wingle(Squamopteromys caudopluma), a rattile of the shingle family, particularly the lineage of arboreal tree shingles. Equipped with scaly plates modified from hair, most shingles use them for armor: using hypertrophied hair erector muscles to raise their plates or clamp them down. This movability, however, allows the wingle to utilize its plates, now thin and papery with a hard central shaft, for a new purpose entirely– spreading them out into a gliding surface, with its tail also adorned with plumes of its own that allow it to change direction. Wingles are omnivores, and use their skill to roam the forest quickly to find a meal: they are not picky eaters, and fruit, fungi, bark, sap, nectar, insects and occasionally smaller rattiles are well on the menu.
However, evolution is ever ongoing, and different species respond differently to changing pressures: forcing them to either become more specialized or remain generalist. With the abundance of ratbats equipped with true flight, the parachu does well staying in such a niche. The wingle, on the other hand, is pressured to migrate further and further by competition from other rattiles diversifying in the Temperocene: and gradually, the modified erector muscles that once just folded and spread their plates developed brief vertical twitches to prolong their glides to access new trees– in ever increasing distances. Soon, in time, the ratbats and pterodents will be joined by new company in the sky ever more crowded and diverse: by something completely unlike anything that had ever flown before.
▪▪▪▪▪▪▪▪▪▪
The Early Temperocene: 140 million years post-establishment

Shroomor Has It: A Lingering Legacy of a Darker Past
Dawn breaks on the steppes of Early Temperocene Mesoterra, as on the plains, the animals of the grassland begin to stir, to begin their day and their usual, instinctive routines like they alwas had for millennia. Among them, in the shade of a small grove of trees, a pair of loupgaroos– now Mesoterra’s top predator since the extinction of their fiercer relative the ripperoo at the end of the Glaciocene– opportunistically stumble upon the decaying, half-eaten remains of a dead podothere. Loupgaroos were normally active predators, going after live prey they hunted themselves. However, they did not hesitate to take advantage of an easy meal, even if it was far from fresh.
Equipped with strong stomachs and hardy immune systems, the podotheres of Mesoterra have a strong resistance to many diseases, thanks to small-gene mutations that protected them from cellular damage and negative gene expressions. This adaptation, ocurring in their ancestors, the drundles, may indeed be the reason why they kept their most bizarre adaptation yet– a rear-shifted ribcage that functionally granted them additional neck vertebrae– without any problems even as they became more active, with such conditions normally being found only in slower-paced mammals.
But not all is perfectly right with this pair of scavenging loupgaroos, as becomes clear as, momentarily startled by ambient sounds, the female raises her head in surprise, surveying the surroundings for danger– and revealing, on her snout, a set of small, telltale growths that may be alarming to one knowledgeable of the horrors of the past.
But, fortunately, the plague of eons ago has passed. Too virulent and destructive for its own good, much like its very hosts, the contagious cancerous strain, known as NO-SIHTT, had destroyed itself, just as its hosts did themselves, by destroying what it depended on to survive until it had nothing left for itself. These growths on the loupgaroo’s face, however, are a different story. It is a far more benign, and far more bizarre strain, that, through eons, has surprisingly found a niche in the environment and become part of the world around it: the shroomors.

From the original strain that infected the long-gone harmsters, a few would jump to closely-related species through passive infection, and end up taking seed in related species: ones genetically similar enough to the harmster as to allow it to thrive. First to be hit was the harmster’s direct ancestors, the ripperoos, which were similarly wiped out, and next to the more-distantly-related loupgaroos, who bear this condition to the present.
But it would be inaccurate to call the shroomors a “disease”, as, except in extremely rare circumstances, it was not at all harmful to its host: it was a benign commensal no more damaging than a few wart-like growths. It spread through scavenging, as loupgaroos ate dead animals as eagerly as they hunted their own meals, but they were kept at bay by the host’s immune system– something that the shroomor exploited in its new, bizarre life cycle.
Travelling on the host, the shroomor was given easy access to find new seeding ground. But instead of infecting living hosts, it grew on dead ones: once it was stimulated by a suitable environment, one without an immune response, it would then begin to grow quickly: forming fleshy, pink cauliflower-like growths very similar to those that adorned the infected harmsters’ faces during the height of the outbreak millions of years long past, the closest one could come to literal walking corpses.
But this time, it would feed upon a decaying, dead host rather than a live one: the enzyme excretions the cancer cells once released to latch onto blood vessels to fuel its growth, now being used like digestive enzymes to break down dead tissue and absorb its nutrients through its cell membranes, allowing it to divide and grow over a span of several days. The entire mass is made solely of undifferentiated cells, so it rapidly divides quickly as long as it has access to nourishment, but eventually, when conditions turn out poorly, as it runs out of nutrients or the carcass begins to dessicate, the shroomor mass slowly dies off, requiring a warm, moist environment to thrive.
But then came a unique innovation: certain cells, on the outside of the mass, became more resistant and could remain dormant for longer periods until conditions were right. As such, these specialized cells became key to the shroomor’s survival, as the rest of the mass died off: they would remain on the carcass until a suitable scavenger came along and ate from it, the loupgaroo now a vector that allows it to be transferred to new growing grounds to flourish elsewhere.
This unusual life cycle, like everything else on the planet, was a unique product of natural selection: more virulent strains of the transmissible tumor have long since died out, with selective pressures favoring those that could live surprisingly amicably with their hosts. While the plague-like infection of NO-SIHTT allowed it to spread quickly, it was a short-term gamble, and the strains that had moved on to other, related host species, that could last on them longer without becoming lethal, were the strains that persisted. Having latched on to scavenging carnivores, it was inevitable that a few cells from the host’s benign warty growths would be dropped off in the carcass, and the hardiest cells that could survive the longest away from a living host persisted, gradually by chance having some actively grow on the carcasses to be passed on by the next scavenger: and slowly some would develop the ability to become more resilient to conditions while they waited for a new host. The ones that remained too active exhausted themselves and died out, the ones that were too sensitive were killed by the unsuitable environment, and what was left were those that grew quickly when conditions were favorable but could also become resilient single cells with thick outer capsules that could conserve their energy by halting cellular processes. Growing on dead decaying matter and spreading itself by what were essentially spores, acting as a decomposer contributing to the breakdown of dead organic matter, the end product was, for all intents and purposes, a mammalian fungus.
This was the harmsters’ final legacy: the disease that destroyed them, borne and cultivated by their own vile ways that ultimately bit them back hard in a terrible, karmic blow, now flourishing in the wild as an animal that could barely even be considered an animal anymore, despite its genetics that would say otherwise. In a way, a part of the harmsters lives on, in a form thankfully devoid of conscious thought or their murderous nature, managing something, in all its simplicity, that its source could not: the ability to find a stable purpose in the natural order, to thrive sustainably in balance with their surroundings– and to finally coexist with other forms of life without being a blight upon them or the rest of the world.

▪▪▪▪▪▪▪▪▪▪▪▪
The Early Temperocene: 140 million years post-establishment

Ocean In Motion: Marine Life of the Early Temperocene
A global increase in temperatures following the end-Glaciocene mass extinction has opened up new ecosystems in the oceans of HP-02017. Indeed, as the ice caps melted in the Temperocene temperatures the sea levels once again have risen, to an extent that coastal regions have flooded into shallow seas, and now as much as 80 percent of the planet’s surface is covered by water.
It is tempting to think of the ocean as merely one big biome, but the truth is far more complex. Here unusual dynamics in the ecosystem allow a far wider range of ecological niches to overlap and coexist, and a tangled food web with numerous trophic levels exist due to the existence of numerous types of producers at its very base, with algae, marine plants and phytoplankton to name a few, which in turn allows a staggering diversity of life to thrive. Floating mats of seagrass, forests of kelp, shallow reefs, open seas and coastal shelves all harbor different, specialized organisms, just as how jungles, tundras, savannahs and deserts would on dry land.
And there was no better age than the Temperocene to showcase marine biodiversity, as a wide range of separate unrelated clades now make a living in the sea: both creatures native to the seas since the beginning, to secondarily-aquatic species descended from numerous hamster lineages: all converging on the abundance of resources and vacant niches that the sea had to offer.

In the shallows, enormous reefs rise above the sea floor like castles, providing homes for a wide diversity of sea life. But, as with nearly everything on this planet, appearances can be decieving, and not everything is what it may seem.
Some of the reefs are comprised of millions of small, hard-shelled barnacle-like creatures rooted in place and extending long feathery appendages to scoop up drifting particles and plankton, while six-limbed starfish-like creatures crawl slowly across the seafloor, foraging for food and feeding upon the shelled reef-creatures. But despite appearances, both of these invertebrates are actually snails: the sessile filter-feeders are quillnobs, hard-shelled snails that have free-swimming larvae, juveniles that settle to the bottom and slither about like typical snails, and as adults finally fuse themselves to one spot and never move again, using modified extensions of their gills to feed. The “starfish”, on the other hand, are asterisks: shell-less gastropods with six extensions of their foot that, equipped with suckers, can anchor themselves onto surfaces with incredible strength.
Meanwhile, drifting through the currents are transluscent, tentacled cnidarians: mockjellies. They are, true to their name, not jellyfish, but neotenic coral larvae that never matured into sessile reef-building adults, becoming what are technically speaking enormous plankton. Some, such as the smaller, stinging stheno, are predators: possessing stinging cells in their tentacles used in capturing small prey such as swimming crustaceans, while their bigger relative, the euryale, have developed a unique symbiosis with photosynthetic algae: allowing them, once adult, to live almost entirely on sunlight alone: but still possessing stingers that ward off most predators from eating them.

In the absence of fish, Earth’s most abundant marine swimmers, the ecological niche was instead filled by the shrish: descended from krill, these shrimplike swimmers dominate oceanic and freshwater habitats alike, in a staggering thousands of species: not quite as diverse as they used to be in the Rodentocene when they were the sole rulers of the sea, but still ever present wherever aquatic ecosystems were: as the long-bodied centipede-like shreels, the bottom-feeding trilobugs, or the predatory shrarks, which once were the top of the food chain, but with the coming of aquatic hamsters have now been relegated to smaller mesopredator roles, with none growing larger than two feet.
The success of the shrish has even brought about an evolutionary trend that occured so frequently on Earth that it would have been more surprising to not happen here: a crustacean with a reduced, folded abdomen, an armored thorax, a rounded body shape and two pincers for feeding–basically, a crab. Known as the shrabs, these amusingly-familiar shrish are bottom-feeding opportunistic omnivores, eating carrion, small animals, algae, corals and even one another. And most remarkable of all are some shrabs that have left the sea entirely to make a living on an entirely new frontier: dry land. These terrestrial shrish are the first of their kind to leave the sea, and are able to survive near-indefinitely on land, as long as their gills remained moist.
But the shrish are not alone in the seas. In the Glaciocene, another marine clade had rose to prominence: the pescopods, a group of swimming sea slugs that propelled themselves with rhythmic undulations of their fin-like foot. In many niches the pescopods have supplanted the shrish thanks to their rasping radula that allowed them to feed on a wide variety of food. However, shrish and pescopods alike have thrived side-by-side thanks to niche partitioning, and due to their widely-disparate anatomy– shrish being armored and exoskeletal and pescopods being soft and rubbery– the predators that feed on them have also evolved dentition suited for one or the other, allowing again for partitioned niches that permit a great diversity of clades in the sea.
Another successful gastropod is the skwoid: a marine gastropod descended from the shelled, tentacled notilus, which is also still very abundant today. Skwoids, however, have ditched their protective shells for an internal support pen, as they are speedy predators suited to chase down their prey with the help of excellent vision from eyes mounted on retractable stalks. To defend themselves in turn from their own enemies, they spray thick mucous secretions to ensnare an attacker: though some skwoids have innovated on this defense and instead became ambush hunters in reefs: spinning mucous webs like some undersea spider for some unlucky shrish or pescopod to blunder into and find itself trapped.

With such a wide range of available resources, the aquatic hamsters of the sea have equally reached enormous diversity. Niche partitioning came to govern their evolution in unusual ways, producing numerous, disparate and completely unrelated species living side-by-side with minimal competition.
The mass extinction at the end of the Glaciocene, partly the work of changing climes and partly the work of the long-gone harmsters, had freed up many marine megafauna niches, and the Temperocene began with a race to fill it from all sides. With so much food available there was enough for everybody so long as they were adapted to consume it: and so marine megafauna hits its peak in the diverse and productive Temperocene seas.
The old rulers of the sea are still here: the seavers. Now, however, they are a fraction of their former glory, though still quite plentiful. Descended from the last Glaciocene survivor, the derelict seaver, they are now much smaller, as warmer seas and fewer cold-water upwellings were no longer conducive for giant filter feeders. Today, the derelicts have rebounded, feeding on swarms of small shrish and zooplankton, but now they never exceed more than eight meters in length.
Cricetaceansandbayvers as a whole still dominate the seas, but they have plenty of new neighbors. Among them are aquatic blubbats from the continent of Peninsulaustra, who, while heading out to sea to feed, came to colonize small island archipelagos in the open ocean and are now a common sight across the Centralic Ocean. They coexist by specializing on different prey: blubbats, with pointed conical teeth, are well-suited for tackling soft-bodied pescopods, while the seal-like bayvers with sharp slicing incisors and crushing molars are well-adapted for shrish, and the long-snouted roddolphs prefer skwoids, learning through social teaching on how to remove the mucous gland and sharp internal pen to make them safe to eat.
Herbivoroushamatees also cruise the warm shallows, with the largest, the whalerus, specialized to eat a floating seagrass known as coast kudzu, which, fittingly, grows at an absurdly fast rate and can form immense masses that blanket the surface. They are also an extremely abundant source of food for whaleruses that they can sustain sizes exceeding ten meters, and gradually shift away from filter feeding to specialize on this resource: an adaptation that may have been a saving grace of the derelict seaver that earlier had declined from their competition.
The rattiles too have taken to the seas, with the monisaurs: some now fully aquatic and bearing their independent young in the sea. The vast majority of them, with a few herbivorous exceptions, are durophages: equipped with broad, blunt teeth and strong jaws, eat mostly hard-shelled invertebrates, with shrabs, quillnobs and bivalves comprising most of their diet.

The monisaurs now are not the only rattiles in the sea: aquatic shingles, known as sterapins, have made their way in too. Their armor-plated terrestrial ancestors being resistant to irritating chemicals from eating land plants and invertebrates with such defenses, the sterapins were well suited to take on something in the ocean that no other animal eats– the mockjellies. Mostly water and armed with stingers, hardly anything else bothers with them, as they are scarcely worth the effort. But where there is a niche something will evolve to fill it: and the sterapins did, uncontested in their unappetizing diet by any other competition, even if they did have to eat plenty of mockjellies just to gain sufficient nutrition.
Another common species in the seas are rodders: small remnant searets of distant relation to the now-gone leviahams that once roamed the seas. They avoid competition in the hectic ocean environment not by specializing as others have done, but by becoming generalists. Thus they are not limited to one food source as competitors come and go, and these small, semi-aquatic omnivores forage both underwater and on the shore, their flexibility being their key to success to survive where their giant cousins could not.
With so many new competition some of the bayvers and cricetaceans have adopted odd new specializations to ease the pressure of sharing their food sources and territories with crowds of newcomers. The whiskered walmus, a species of hamatee, has returned to a mostly-carnivorous bottom-feeding diet, probing out the bottom substrate for buried invertebrates such as clams, worms and shrabs. Meanwhile, the porpoid, a cricetacean relative of the roddolph, relieves competition with its long-snouted kin by specializing on shelled notiluses, aided by rounded, globe-shaped teeth that can easily crack the shells to get the soft meat inside.
One of the more unusual hunters in the sea, however, comes not from the water, but from the sky. The pterodents, a young but very promising clade, too have made a foray into the blue depths, in the form of the wanderganders: sea-adapted soaring flyers that, in just five million years, have radiated into over a dozen species, comprised of surface-skimmers, plunge-divers, shore-probers and even sky-pirates that harass smaller pterodents and ratbats in order to steal their food.

With such a vast diversity of large prey species it was no surprise that marine predators too would experience a boom of diversity themselves with so many different prey to hunt. These were the phorcas, cricetaceans adapted to tackle large prey, and with the disappearance of their old rivals, the leviahams, at the Glaciocene-Temperocene boundary, they would quickly fill the role of top marine predator left in the leviahams’ wake, now spanning over twenty distinct species: all adapted for taking on different game.
Some, such as the sheartooths, specialized on small, fast prey, like blubbats and small porpoids and blippers, which they pursued in short, quick bursts and dispatched with conical stabbing teeth, primarily hunting quarry that could be consumed in a bite or two. Others, such as the whillers, went after larger prey, coordinating in social groups with the help of distinct dark-and-light contrasting markings for communication, allowing them to fell large whaleruses and seavers. Still others adapted to armored, slow-moving prey like sterapins and monisaurs, like the striped shellbanes, with broad pointed teeth with fewer cusps designed to concentrate their bite force onto small areas and penetrate even thick rattile scales.
With many predators abound and even more prey, death was always around the corner and carcasses were not hard to find: either by natural causes or the leftovers of predatory phorcas. As such, a specialized scavenger, the seayena, would diverge from the shellbanes, adapting to feed upon bones to access the marrow. Broad crushing molars and the strongest bite force of any living animal on the planet allow the seayena to exploit a resource others could not: consuming not only the marrow but the bones themselves, which powerful digestive juices can handle perfectly well to extract the maximum nourishment from their unsavory diet.

Predatory phorcas have become so abundant and numerous in the Temperocene that they now occupy virtually all the ranges in the oceans worldwide, sustained by the vast abundance and diversity of megafaunal prey that flourish in the seas. This has reached such an extent that an entirely new ecological niche would open up in the marine ecosystem: a predator that specializes to prey on phorcas themselves.
Enter the sarchon(Phorciphagus tyrannicus): ten meters in length, it is unquestionably the top rung of the Temperocene marine food web, and in fact is on an entirely new level of its own– a top predator specialized to eat other top predators. Its diet, based on local availability, too includes other, more placid prey such as seavers and whaleruses, and likely began its predation of its fellow phorcas from initially just killing them to dispose of competition, before finding them an equally nutritious and abundant food source in their own right. As such the sarchon has attained truly impressive weaponry: rather than the sharp-cusped molars of more-typical phorcas, it relies on its incisors, ever growing like those of a more typical rodent, as flesh-slicing shears, with its first molars modified into broad cutting edges and its second and third molars gone entirely. Its head is heavily armored with thick, keratinized plates to protect its vulnerable snout from the bites of smaller phorcas, leaving them defenseless while the sarchon deals its lethal blow.
Unchallenged by virtually anything else in the sea, with no natural enemies of its own to fear, the sarchon spends most of its time lethargically floating near the surface to rest and digest, and, contrary to its fearsome feeding strategies, is otherwise a very lazy creature: like all cricetaceans, it only rests part of its brain at a time to breathe at the surface, and can spend as much as twenty hours a day just drifting around in a half-asleep stupor, occasionally floating belly-up at the surface to let sea ratbats pick parasitic louse-like shrish off its skin. Subsisting off such a nourishing diet, and without any danger to concern it, the sarchon hunts only once every few days, sometimes as little as once a week after a large meal, and spends much of its free time in a surprisingly relaxed state: its epic hunts towards other predators, which have come to define it, actually taking up very little time in the long, fascinating and complex life of the Temperocene’s ultimate ocean carnivore.
▪▪▪▪▪▪▪▪▪


I haven’t been posting here nearly as much as I use to. I tend to post more regularly on Instagram or DA so if you are interested head over there to see my work more regularly.
some alien critters because I needed a break from all the demons.
Post link


Almost Real Vol. 5 has reached all its stretch goal tiers and has THREE DAYS LEFT before its campaign ends Thursday 6 • 16 at 9AM PST!
Free itslittletunny sticker sheets, 6x9″ prints of our cover critter’s patch design, and artist bonuses for all: what’s better than this?!

Check us out and pledge today before time runs out!
Post link



Almost Real: A Speculative Biology Zine’s 5th Volume all about mythology has passed its second stretch goal!
Thanks to your help and pledges, every backer at the “CONFOUNDING CREATURES” tier and up will have a FREE complimentary 8.5x11″ sticker sheet of our issue’s entries drawn by campaign guest artist @itslittletunny!

Pledge now to receive your rewards and carry us further into our campaign before time runs out!
Post link


Barely over 24 hours and Almost Real is sitting at over 50% FUNDED with 200+ backers!

We can’t thank you enough for your support already! ✨Check us out today to see our campaign previews, artist roster, stretch goals, and MORE: only on Zoop.gg!

Post link
 is now LIVE on Zoop.gg! ✨Legends new")
✨Almost Real: A Speculative Biology Zine Vol. 5 (BIOTECHNOLOGY) is now LIVE on Zoop.gg! ✨
Legends new and old come to life in Almost Real’s fifth mythological volume, powered by our contributors’ speculative imaginations.

Click the links to pledge today before our campaign ends on June 16th!
Post link
 launches May 16th on ZOOP! ✨Legends ol")
✨Almost Real: A Speculative Biology Zine (Vol. 5 - MYTHOLOGY) launches May 16th on ZOOP! ✨
Legends old and new come to life with Almost Real’s fifth volume; bringing a dozen speculative entries examining all sorts of fantastical creatures!

Head to Twitter to see the full announcement thread and extended sneak peeks from the @FortunaMedias account, but for now: mark your calendars and save the date for Almost Real’s 5th installment on the Kickstarter-alternative platform Zoop: a crowdfunding site dedicated entirely to comics and graphic novels (without the pledge to blockchain)!
Our contributors for Volume 5:
- Laura Wilson (twitter: @LauraMakesArt)
- Sophie Morse | SOPHARIUM (twitter: @Sopharium)
- October Seagrave (twitter: @Zoophagous)
- KoryBing (twitter: @KoryBing)
- Bree Paulsen (twitter: @breebird33)
- CrabdominalPain (twitter: @CrabdominalPain)
- Shirley Jackson|LIONINTHETREES (twitter: @LionintheTrees)
- E. “Papermonkey” Jokinen (twitter: @Paperiapina)
- Genystar(twitter: @GGenystar)
- GEN H. | PAPAYAYA (twitter: @_papayaya)
- Lonnie Garcia (twitter: @Sixgilis)
- Cy (twitter: @TheCydork)
- Iben Krutt (twitter: @IbenKrutt) (cover artist)
- Mar Spragge (twitter: @ParasiteBeans) & Puzzled Jay Productions (insert poster)
We’ll see you May 16th with another legendary volume of Almost Real: A Speculative Biology Zine!

Post link

Almost Real’s 4th Volume, all about Biotechnology, has come to a close!

We ended on an incredible three stretch goals broken (with two of those being well-deserved artist bonuses for all involved contributors!) None of this would’ve been possible without our artist’s imaginations and our backer’s support, and thanks again for helping us make Almost Real… real!
Check out the Kickstarter update here with more info, watch for backer surveys going out in mid-May, and we’ll see you soon!

Post link

DAWN OF THE FINAL DAY For Almost Real’s 4th Volume- All about BIOTECHNOLOGY!
Back us before time runs out (and to see if we can’t break our 3rd stretch goal of a 2nd Artist Bonus for all contributors)!

Post link
 is ENTERING ITS FINAL HOURS! Sticker S")
Almost Real: A Speculative Biology Zine VOL 4 (BIOTECHNOLOGY) is ENTERING ITS FINAL HOURS! Sticker Sheet Stretch Goal Unlocked! ✨
Ending on Monday April 19th at 12PM (PDT), we’ll be closing out Almost Real’s 4th Kickstarter! Since our last update, we broke our 2nd stretch goal of free 8.5x11" sticker sheets illustrated by V Swain if you’re a backer of the SCANNED BLUEPRINTS tier and up!

As it says on the battery, our next stretch goal is our 2nd artist bonus for contributors at $24,000! We’d be more than thrilled at the idea of paying our contributors as much as we can, so get your last minute pledges and/or adjustments in before Monday.
Head here to support the 4th installment of this spec bio collection before the Kickstarter campaign ends!
Post link

FINAL WEEK for Almost Real Vol. 4! First Stretch Goal Reached!
Our Kickstarter has officially started its last week, formally ending on April 19th at 12PM PDT! Thank you all so much for your rousing support to put us at our goal already- your pledges mean the world to us creating this corner of spec bio, and we hope you enjoy what biotechnological wonders our contributors have to offer.
In other exciting news: in the wee hours of the morning, we hit our first stretch goal of $19,500, which means artist bonuses for all our contributors!

Our next goal will be at $21,000, in which we’ll have V Swain illustrate us an 8.5"x11" sticker sheet featuring every design from the entry! And after that? Another artist bonus at $24,000, so let’s see how much we can crank out in our final week!

✨Head here to back Almost real’s 4th volume, all about BIOTECHNOLOGYtoday! ✨
Post link
 is a Project We Love on Kickstarter,")
✨Almost Real: A Speculative Biology Zine Vol. 4 (BIOTECHNOLOGY) is a Project We Love on Kickstarter, and has only $7,000 left to go before funding!
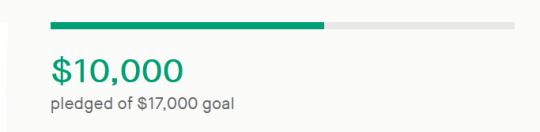
Wow. Truly, thank you all for an INCREDIBLE first couple of days with your support! With this influx, we’ll be funded in no time, and we hope to crack as many stretch goals (and artist bonuses!) as possible before April 19th!
Head over to our Kickstarter page updates to see a special sneak peek from contributor Mike McGhee’s entry, and pledge today and help make Almost Real… real!

Post link
 is now LIVE on Kickstarter! The Alm")
✨Almost Real: A Speculative Biology Zine Vol. 4 (BIOTECHNOLOGY) is now LIVE on Kickstarter!

The Almost Real series is back with Volume 4: powered by Biotechnology, and inspired by the imaginations of over a dozen contributors.

Click the links to pledge today before our campaign ends on April 19th!
Post link
 launches March 20th on Kickstarte")
✨Almost Real: A Speculative Biology Zine (Vol. 4 - BIOTECHNOLOGY) launches March 20th on Kickstarter! ✨
Pull up a chair, dust off those blueprints, and see what specimens you’re working with as Almost Real’s 4th Volume takes you on a journey through biotech of all shapes and sizes.

Head to Twitter to see the full announcement threadand extended sneak peeks from the @FortunaMedias account, but for now: mark your calendars and save the date for Almost Real’s 4th installment!
Our full contributor list for Volume 4:
- Rowena V (twitter: @ProfessorDaft)
- Iris Jay
- Moa (Avian King) (twitter: @Moafowl)
- Roxy Valdez (twitter: @BigHugeFrog)
- Blue Delliquanti
- MechanizedMonsters
- Takamatsu-kun (twitter: @Takamatsu_kun)
- Galen “Strop” Haecky
- Jack Friedman (caba-111)
- Mike McGhee(site, instagram: @mmmmmike_mcgheeeee)
- Hotkoin (instagram: @Hotkoin)
- Ryan McCowan (twitter: @RyanResolute92)
- Emely Pascual (twitter: @Aquiboni) (cover artist)
- Hana Chatani (instagram: @hanachatani) (insert poster)

We’ll see you March 20th with another spec-bio filled volume of Almost Real: A Speculative Biology Zine!
Post link